转录
一、转录的一般特点
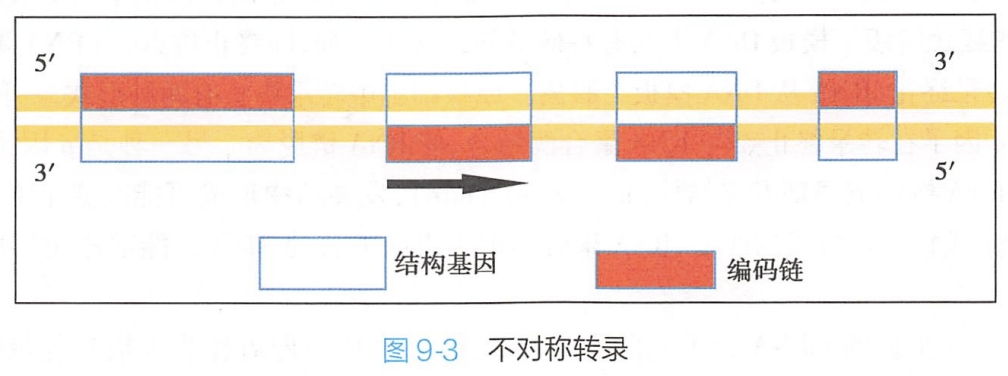
DNA链是基因转录的模板。在双链DNA中,作为转录模板的链称为模板链(template strand)或反义链(antisense strand),即与mRNA互补的DNA链;与模板链互补的另一条链称为编码链(coding strand)或有意义链(sense strand),该链与转录产物的序列相同,只是在转录中将DNA中的T变为RNA中的U。
模板链并非总在同一单链上,如图9-3所示,在DNA双链的某一区段,以其中一条单链为模板,而在另一区段,以其相对应的互补链为模板。这种DNA链的选择性转录也称为不对称转录(asymmetric transcription)。
转录的本质是一个以DNA双螺旋链中反义链为模板,以四种核苷三磷酸ATP、GTP、CTP、UTP为原料,在RNA聚合酶作用下,遵循碱基互补配对原则合成RNA的过程。
RNA合成的方向是5'→3',生成的RNA链与模板链反向平行,游离的NTP(Nucleoside triphosphate,核苷三磷酸,与dNTP相对)只能连接到RNA链的3'-OH端。
在RNA合成中不需要引物。
原核细胞和真核细胞区别:
- mRNA:原核细胞转录形成的mRNA分子一般不需要加工,在合成后即可作为模板参与蛋白质的合成。在真核细胞中,转录生成的RNA是初级转录产物,必须经过不同方式的加工和修饰才具有生物活性。
- rRNA:原核细胞需要在合成初级转录产物的基础上,进一步剪切修饰才能成为具有生物功能的成熟分子。例如,rRNA基因的初级转录产物在核酸酶的作用下(真核是核酶),剪切成3种rRNA,即5S rRNA、16S rRNA、23S rRNA,三者比例为1:1:1。
- tRNA:原核生物tRNA成熟的机制与真核生物相同。
二、原核细胞转录
(一)RNA聚合酶和ρ因子
RNA聚合酶和ρ因子是原核细胞基因转录的主要因子(应该不算转录因子)
1.RNA聚合酶
原核细胞中只有一种RNA聚合酶,催化转录形成各种RNA分子。大肠杆菌的 RNA聚合酶是由4种不同亚基构成的复合体,记为α2ββ'σ,称为全酶。没有σ亚基的复合体称为核心酶。
σ亚基的主要功能是识别原核细胞基因上游的启动子(识别区),启动基因转录。核心酶的作用是催化RNA的聚合。其中β'亚基能与DNA模板结合,β亚基能与核苷三磷酸结合,并在磷酸二酯键形成中起作用。
2.ρ因子
ρ因子是一种蛋白质。当转录进行到基因末端时,ρ因子能识别终止信号并与之结合,使RNA聚合酶不能继续作用而使转录终止。
(二)mRNA
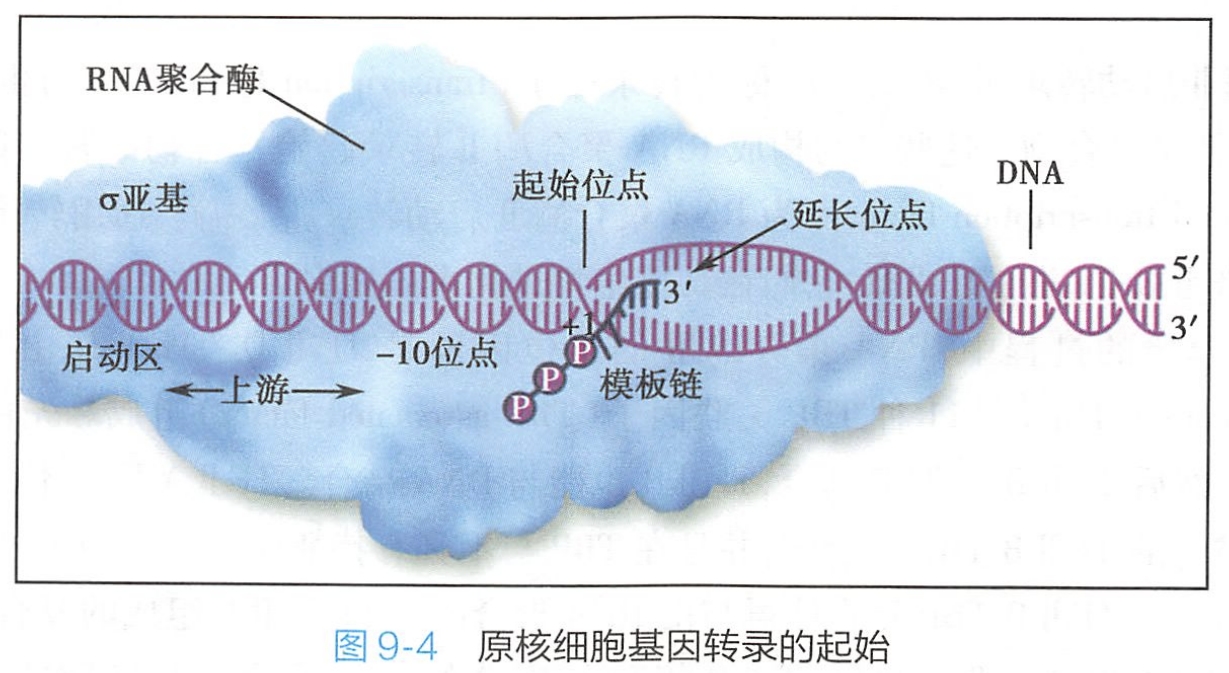
1.起始阶段
首先RNA聚合酶的σ亚基识别基因上游的启动子识别位点,使全酶与启动子结合并形成复合体,这是转录起始的关键,随后DNA双链从局部被打开,以反义链为模板,按照碱基互补配对原则将第一个核苷三磷酸结合到模板的转录起始点上,并与第二个核苷三磷酸形成第一个磷酸二酯键。通常,当RNA链达8~9个核苷酸后,σ亚基便从全酶解离出来,至此完成转录的起始(图9-4)。
2.延长阶段
核心酶沿模板链3'→5'方向移动,在滑行中使DNA双链不断解旋,并按模板的碱基序列配对加入相应的核苷三磷酸(ATP、GTP、CTP、UTP),与DNA分子中双链间碱基配对有所不同的是转录中由新加入的U与模板中的A配对,使RNA链以5'→3'方向不断延长。
随着RNA链的延长,核心酶前面的DNA链不断解旋,而核心酶后面已完成转录的区段则重新形成双螺旋。
3.终止阶段
模板DNA上具有特殊的转录终止序列,即终止位点,当RNA聚合酶移动至终止部位时,转录终止,RNA从DNA模板上脱离。原核细胞中转录终止有两种形式,
- 一种是ρ因子依赖性终止,即ρ因子在转录终止点与RNA聚合酶结合,使RNA链脱离。
- 另一种是ρ因子非依赖性终止,由新合成RNA链形成局部发夹结构(hairpin structure),发夹结构形成可能改变了RNA聚合酶的构象,导致了酶-模板结合方式的改变,RNA聚合酶则不再向下移动,磷酸二酯键停止形成,RNA从模板脱离。
三、真核细胞转录
真核细胞中含有多种RNA聚合酶(表9-1)。

(一)mRNA
真核细胞mRNA成熟前经历首尾加工和中间剪接
1.hnRNA
mRNA是RNA中唯一具有编码蛋白质功能的RNA分子,其前体是编码基因在RNA聚合酶Ⅱ催化下转录形成的。由于RNA前体分子的大小各不相同,被称为不均一核RNA(heterogeneous nuclear RNA,hnRNA),hnRNA需经过剪切修饰才能成为成熟的mRNA。
RNA聚合酶Ⅱ启动转录时,需要一些称为转录因子(transcription factor,TF)的蛋白质参与,才能形成具有活性的转录复合物。这些参与构成RNA聚合酶Ⅱ转录起始复合物的转录因子通常称为通用转录因子(general transcription factor),为RNA聚合酶Ⅱ启动转录所必需。
通用转录因子有TFⅡA、 TFⅡB、TFⅡD、TFⅡE、TFⅡF、TFⅡH,它们在真核生物进化中高度保守。
在基因转录起始的过程中,RNA聚合酶Ⅱ的定位因子是TFⅡD。TFⅡD由TATA-结合蛋白(TATA-binding protein,TBP)和11种TBP关联因子(TBP-associated factor)组成。
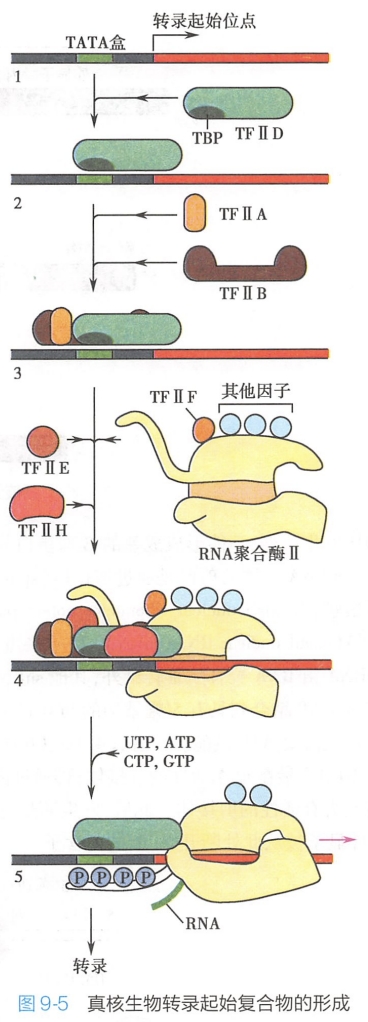
- 先由TBP结合到启动子的TATA盒
- 然后TFⅡB与TBP结合,TFⅡB也能与DNA结合。
- TFⅡA虽然不是必需的,但它能稳定已与DNA结合的TFⅡB-TBP复合体,并且在TBP与不具有特征序列的启动子结合(这种结合较弱)时发挥重要作用。
- TFⅡB-TBP复合体再与由RNA聚合酶Ⅱ和TFⅡF组成的复合体结合。TFⅡF的作用是通过和RNA聚合酶Ⅱ一起与TFⅡB相互作用,降低RNA聚合酶Ⅱ与DNA的非特异部位的结合,来协助RNA聚合酶Ⅱ靶向结合启动子。
- 最后是TFⅡE和TFⅡH加入,形成闭合复合体,完成转录起始复合物的装配。TFⅡH具有解旋酶活性,能使转录起始点附近的DNA双螺旋解开,使闭合复合体成为开放复合体,启动转录(图9-5)。
- 当合成一段60~70bp的RNA后,TFⅡE和TFⅡH释放,RNA聚合酶Ⅱ进入转录延长期。一旦RNA合成结束,转录即终止。
- 有关真核生物转录终止的机制目前尚不清楚。(在基因那一节你可不是这么说的)
2.hnRNA加工
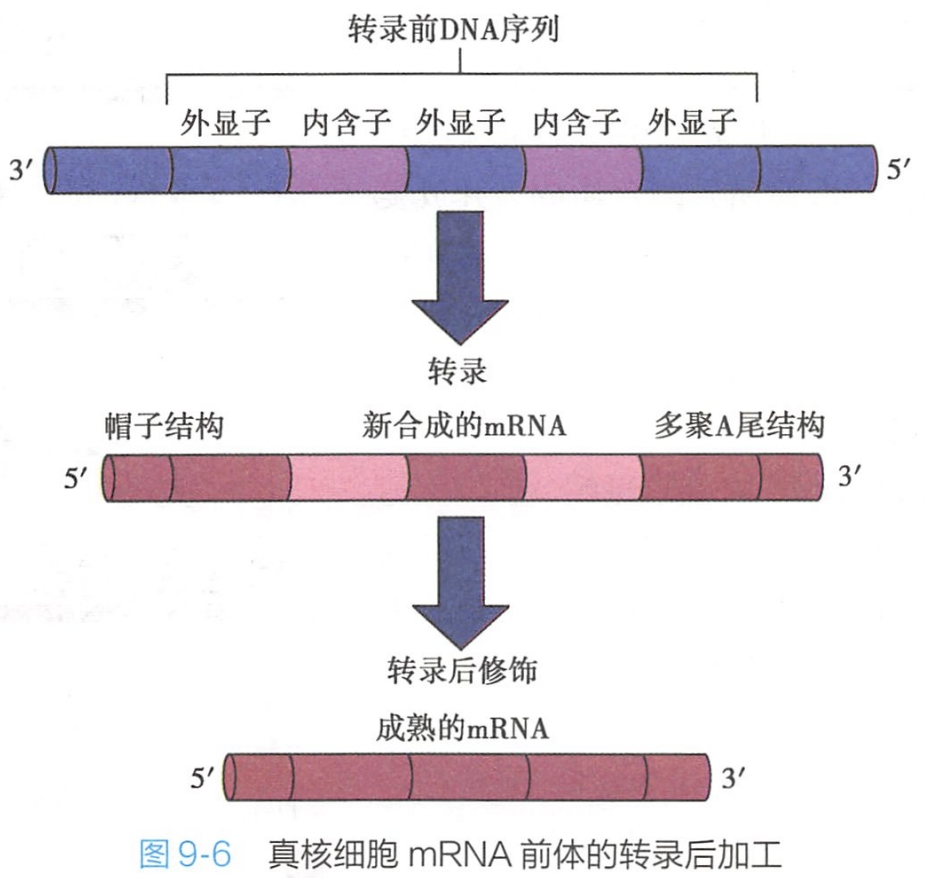
hnRNA加工过程包括戴帽、加尾和剪接(图9-6)。
(1)戴帽
戴帽(capping)是指对hnRNA5'端进行化学修饰,新生的mRNA的长度达25~30bp时,就开始进行戴帽修饰。即:
- 首先在mRNA前体5'端开始的第一个核苷酸上接上一个三磷酸鸟嘌呤。
- 然后在甲基转移酶的作用下,在鸟嘌呤第7位的氨上进行甲基化,形成一个7-甲基鸟嘌呤三磷酸(m7Gppp)的帽子结构。
- 同时在原来第一个核苷酸的2'-O上也进行甲基化,因此,一个帽带有二个甲基。
mRNA戴帽的作用:
- 一是能封闭mRNA5'端,使其不再加接核苷酸,
- 同时也防止转运时被核酸酶水解,增强RNA的稳定性;
- 二是帽子结构能被核糖体小亚基识别,有利于mRNA最初翻译的准确性。
(2)加尾
加尾(tailing)是指对hnRNA3'端的修饰过程,即在腺苷酸聚合酶的作用下,在3'端加上由200~250个腺苷酸组成的多聚腺苷酸(poly-A)的尾巴。
加尾的作用:
- 一方面可使mRNA3'端稳定,防止被核酸酶水解,
- 另一方面有利于mRNA由核到细胞质的转运。
目前发现,决定mRNA前体分子加尾的信号序列存在于RNA3'端,即在发生多聚腺苷酸化位点的上游10~30bp处存在一高度保守的6核苷酸序列(在哺乳动物中为AAUAAA),在多聚腺苷酸化位点的下游常存在富含G和U的序列。这些序列可被含有腺苷酸聚合酶的复合物所识别,进行加尾。
(3)剪接
基因转录过程是以一段连续的DNA序列为模板进行的,在初级转录产物中包含有内含子和外显子序列,在形成成熟mRNA过程中,需将hnRNA中的内含子切除,形成由连续编码序列组成的mRNA模板。
初级转录产物中的内含子常以GU开始,以AG结束,被认为是真核生物基因特有的剪接信号,也称剪接点。几乎所有真核生物基因的内含子均遵循这一GU-AG规则,表明这类内含子存在着共同的剪接机制。
完成hnRNA剪接需要有三个必需的序列:5'GU序列,3'AG序列和分支点(branch site)。分支点位于内含子3'端上游约30个碱基处,为一高度保守的A,在3'剪接位点的识别中起重要作用。
mRNA前体的剪接是通过剪接体(spliceosome)完成的。剪接体大小为60S,由数种小分子核糖核蛋白颗粒(small nuclear ribonucleoprotein particle,snRNP)组成。snRNP由细胞核中存在的一类小核RNA(small nuclear RNA,snRNA)和一些蛋白质组成,常见的snRNA有U1、U2、U4、U5和U6,除U6 snRNA由RNA聚合酶Ⅲ转录外,其他snRNA均由RNA聚合酶Ⅱ催化合成。
剪接过程见图9-7
- 首先 U1 snRNP结合到具有5'帽结构的hnRNA内含子5'剪接点,随后U2 snRNP结合到内含子分支点,这一过程需要ATP供能。
- 接着,U4、U5、U6 snRNP以复合体的形式与先期已经结合于mRNA前体分子的U1、U2装配成无活性的剪接体;
- 再通过内部的重排,U1、U4被排出剪接体,从而使无活性的剪接体转变为有活性的剪接体。
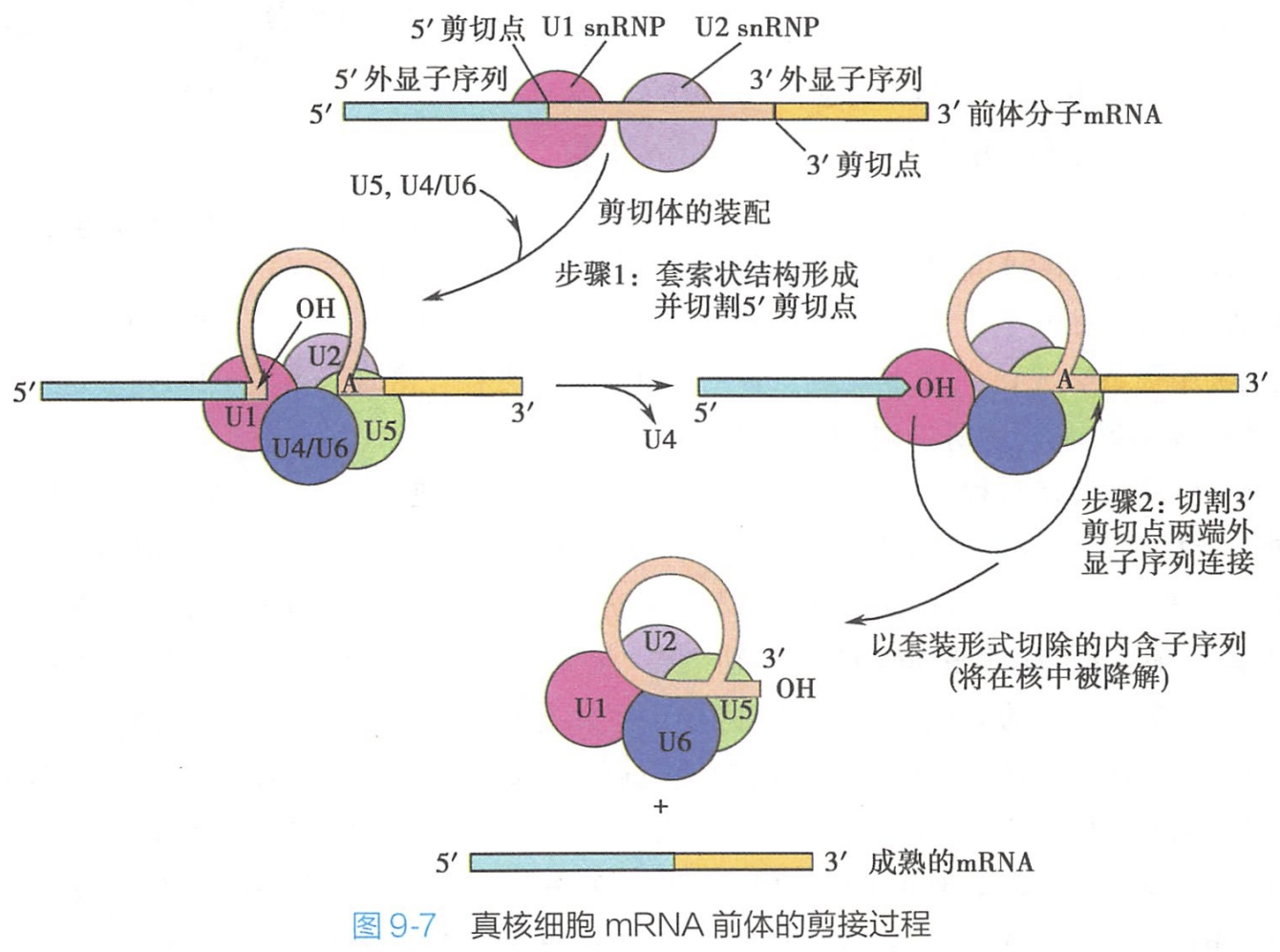
- 此后,发生两步反应,第一步,分支点上的核苷酸A接近5'剪接点,内含子与 5'外显子被从此处断开,切断的内含子5'端与核苷酸A共价连接形成套索状(lariat)结构。
- 第二步,5'外显子上的3'-OH端与3'外显子的起始部位结合,并切割3'剪接点,3'和5'端外显子彼此连接,剪接体各组分和套索状结构脱离,剪接完成。
(二)rRNA
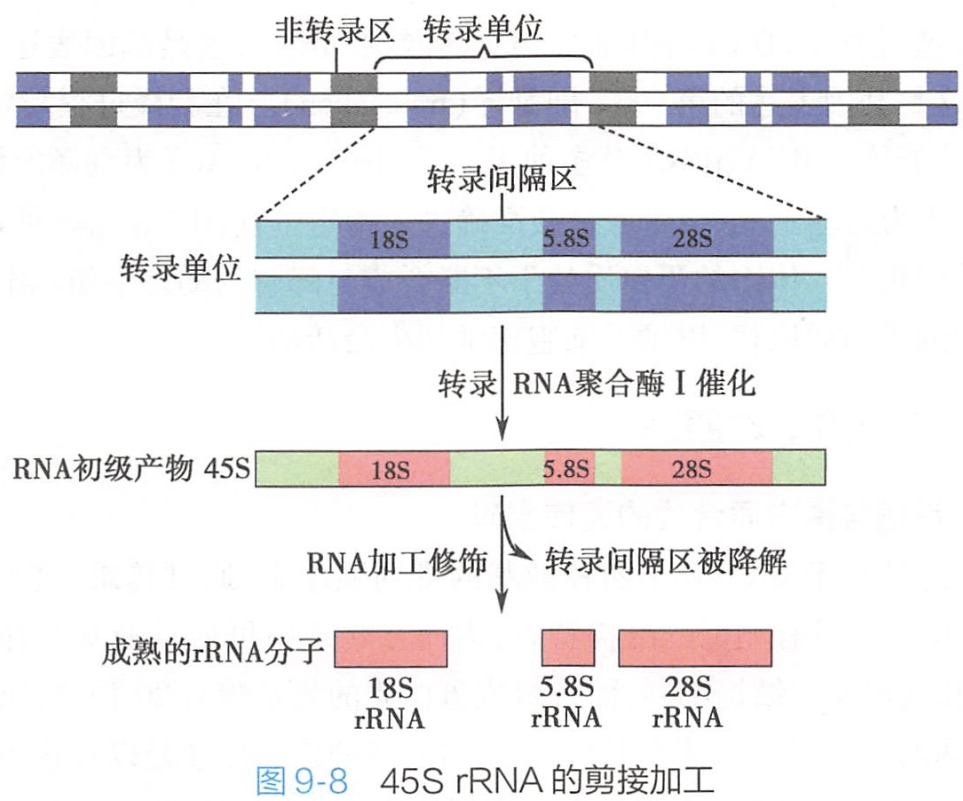
真核细胞中的rRNA基因串联排列于特定的核仁染色质区段,为中度重复序列,每个基因之间由不转录的间隔DNA分隔,这种间隔长度在不同种属生物间差别较大。
每个基因由3个外显子和2个内含子组成,3个外显子依次为编码18S rRNA、 5.8S rRNA、28S rRNA的前体序列,共同组成一个转录单位。在RNA聚合酶I催化下转录形成原始rRNA前体45S rRNA,最终剪切为28S、18S和5.8S rRNA(图9-8)。( #? 你的观点我很认同,但你之前外显子内含子可不是这么定义的)
5S rRNA来自核仁外。5S rRNA是一类特殊的rRNA分子。与其他类型的rRNA分子不同,5S rRNA是由核仁外的基因编码,5S rDNA为串联排列的多拷贝基因,在RNA聚合酶Ⅲ的作用下,5S rDNA转录为5S rRNA。由于在5S rDNA中无内含子存在,所以,由5S rDNA转录形成的5S rRNA无需进一步的剪切加工,即可转运至核仁中,直接参与核糖体大亚基的组装。
rRNA前体的另一种加工形式是甲基化,主要发生在核糖的2'-羟基,一般认为,这种化学修饰有利于前体rRNA的有效裂解。
有研究表明,rRNA的剪接不需要任何蛋白质的参与即可发生,进行的是自身剪接。前体rRNA释放出一种能催化寡核苷酸底物剪接的短链RNA(L19RNA),即为一种具有酶活性的核酶(ribozyme)。
(三)tRNA
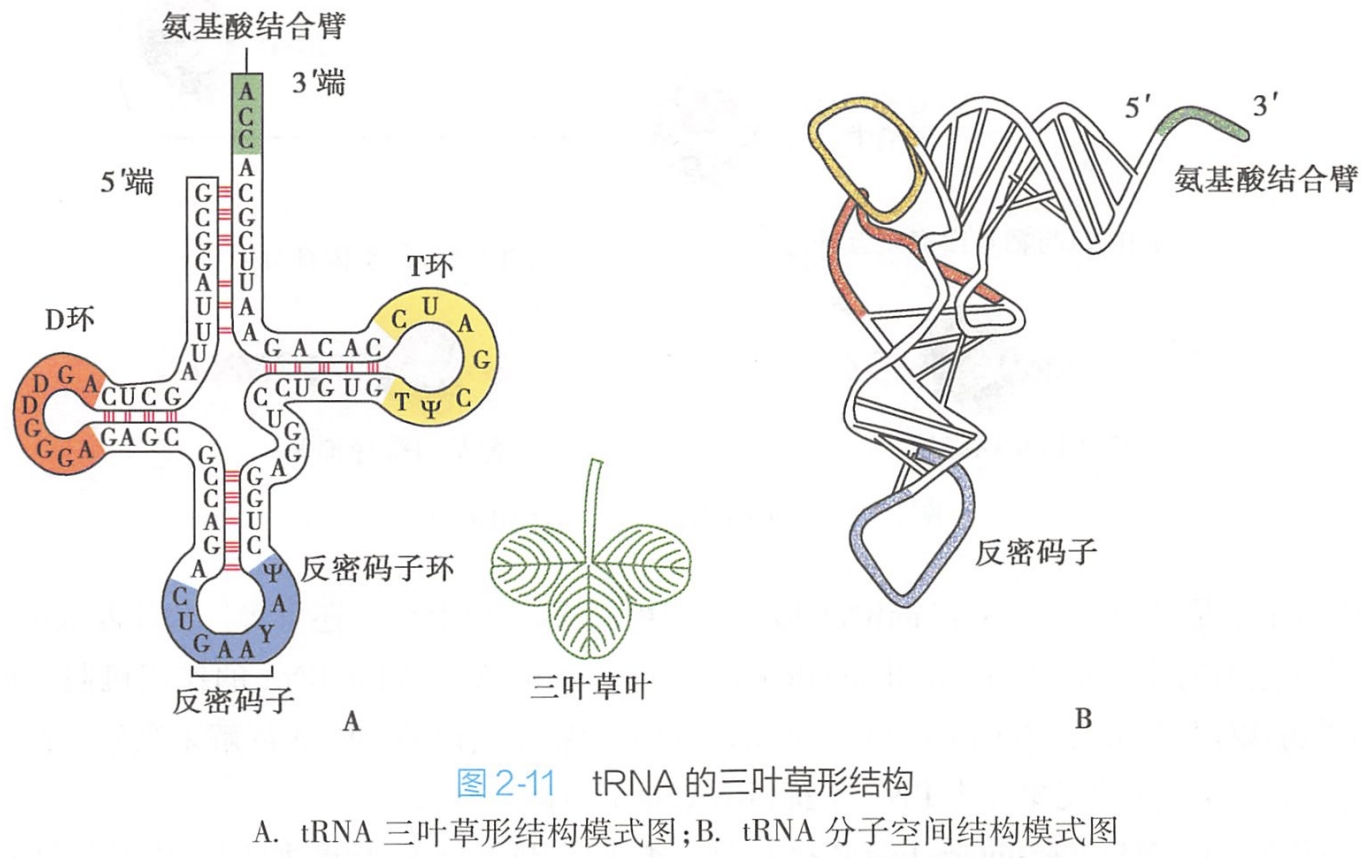
tRNA是一类小分子量的RNA分子。它们能够识别mRNA中的密码子并携带由密码子所指定的氨基酸进行蛋白质的合成。
在真核细胞中含有多个编码tRNA的基因,人体细胞中有1300个拷贝,成簇存在并被间隔区分开,在RNA聚合酶Ⅲ的作用下被转录为tRNA前体。
tRNA前体基因转录需要两种转录因子,TFⅢB和TFⅢC,它们可与tRNA基因转录起始点下游+10~+60中的两个特殊区段结合形成复合物,该复合物与RNA聚合酶Ⅲ结合,启动tRNA基因的转录。真核生物tRNA前体约为100个核苷酸长度,这种新转录生成的tRNA前体一般无生物活性,需要进行剪接和碱基修饰才能形成成熟的tRNA。
1.tRNA前体的剪接
由真核细胞RNA聚合酶Ⅲ催化产生的tRNA前体,在RNA酶作用下,tRNA前体的5'末端和相当于反密码环的区域分别被切除一定长度的多核苷酸链,再由连接酶催化而拼接形成成熟的tRNA。
2.tRNA前体的化学修饰
成熟的tRNA分子中有许多的稀有碱基(rare base),这些稀有碱基常通过化学修饰而成,这些化学反应包括:
- 甲基化反应,tRNA在甲基转移酶催化下,某些嘌呤生成甲基嘌呤如A→Am,G→Gm;
- 还原反应,尿嘧啶还原为双氢尿嘧啶;
- 核苷内的转位反应,尿嘧啶核苷转变为假尿嘧啶核苷;
- 脱氨基反应,部分腺苷酸脱氨基成为次黄嘌呤核苷酸。
3.3'末端加上CCA
在核苷酸转移酶的作用下,3'末端除去个别碱基后,换上tRNA分子统一的CCA-OH末端,完成tRNA分子中的氨基酸臂结构。